
![]()
A blog maintained by Tevita Kete, PGR Officer
Secretariat of the Pacific Community (SPC), Suva, Fiji Islands
This weblog documents the activities of Pacific Agricultural Genetic Resources Network (PAPGREN), along with other information on plant genetic resources (PGR) in the Pacific.
The myriad varieties found within cultivated plants are fundamental to the present and future productivity of agriculture. PAPGREN, which is coordinated by the Land Resources Division of the Secretariat of the Pacific Community (SPC), helps Pacific countries and territories to conserve their crop genetic diversity sustainably, with technical assistance from the Bioversity International (BI) and support from NZAID and ACIAR.
SPC also hosts the Centre of Pacific Crops and Trees (CEPaCT). The CEPaCT maintains regional in vitro collections of crops important to the Pacific and carries out research on tissue culture technology. The CEPaCT Adviser is Dr Mary Taylor (MaryT@spc.int), the CEPaCT Curator is Ms Valerie Tuia (ValerieT@spc.int).

PAPGREN coordination and support
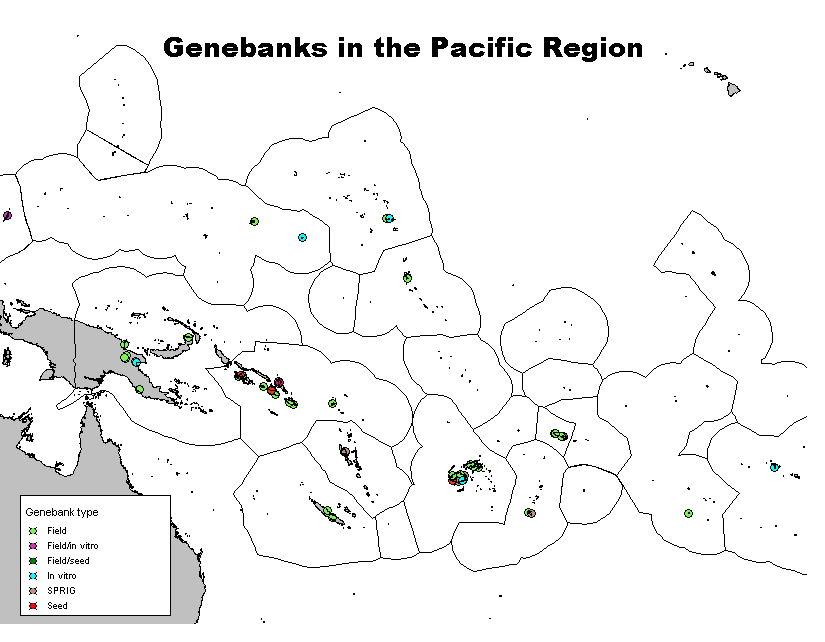
Click on the thumbnail to see a map of the locations of Pacific genebanks. Click here to download a regional directory of genebanks in the Pacific, including information on their location, contact details and holdings.
PAPGREN partners
Mr William Wigmore
Director of Research
Ministry of Agriculture
Department of Resources & Development
P.O. Box 96
Rarotonga
Cook Islands
Tel: (682) 28711-29720
Fax: (682) 21881
Email: cimoa@oyster.net.ck
Mr Adelino S. Lorens
Chief
Agriculture Pohnpei
Office of Economic Affairs
P.O. Box 1028
Kolonia
Pohnpei 96941
Federated States of Micronesia
Tel: (691) 3202400
Fax: (691) 3202127
Email: pniagriculture@mail.fm
Dr Lois Englberger
Island Food Community of Pohnpei
Research Advisor
P.O. Box 2299
Kolonia
Pohnpei 96941
Federated States of Micronesia
Email: nutrition@mail.fm
Mr Apisai Ucuboi
Director of Research
Ministry of Agriculture, Fisheries & Forest
Koronivia Research Station
P.O. Box 77
Nausori
Fiji Islands
Tel: (679) 3477044
Fax: (679) 3477546-400262
Email: apisainu@yahoo.com
Dr Maurice Wong
Service du Developpement Rural
B.P. 100
Papeete
Tahiti 98713
French Polynesia
Tel: (689) 42 81 44
Fax: (689) 42 08 31
Email: maurice.wong@rural.gov.pf
Mr Tianeti Beenna Ioane
Head, Research Section
Division of Agriculture
Ministry of Environment, Lands and Agricultural Development
P.O. Box 267
Tarawa
Kiribati
Tel: (686) 28096-28108-28080
Fax: (686) 28121
Email : agriculture@tskl.net.ki; Beenna_ti@yahoo.com
Mr Frederick Muller
Secretary
Ministry of Resources & Development
P.O. Box 1727
Majuro 96960
Marshall Islands
Tel: (692) 6253206
Fax: (692) 6257471
Email: rndsec@ntamar.net
Mr Herman Francisco
Director
Bureau of Agriculture
Ministry of Resources & Development
P.O. Box 460
Koror 96940
Palau
Tel: (680) 4881517
Fax: (680) 4881725
Email: bnrd@pnccwg.palaunet.com
Ms Rosa Kambuou
Principal Scientist PGR
NARI Dry Lowlands Programme
Laloki Agricultural Research Station
P.O. Box 1828
Boroko
National Capital District
Papua New Guinea
Tel: (675) 3235511
Fax: (675) 3234733
Email: kambuou@global.net.pg
Ms Laisene Samuelu
Principal Crop Development Officer
Crops Division
Ministry of Agriculture, Forests, Fisheries & Meteorology
P.O. Box 1874
Apia
Samoa
Tel: (685) 23416-20605
Fax: (685) 20607-23996
Email: lsamuelu@lesamoa.net
Mr Jimi Saelea
Director of Research
Department of Agriculture and Livestock
P.O. Box G13
Honiara
Guadalcanal
Solomon Islands
Tel: (677) 27987
Mr Tony Jansen
Planting Materials Network
Kastom Gaden Association
Burns Creek, Honiara
P.O. Box 742
Honiara
Solomon Islands
Tel: (677) 39551
Email: kastomgaden@solomon.com.sb
Mr Finao Pole
Head of Research
Ministry of Agriculture & Forests
P.O. Box 14
Nuku'alofa
Tonga
Tel: (676) 23038
Fax: (676) 24271
Email: thaangana@hotmail.com
Mr Frazer Bule Lehi
Head of Research
Department of Agriculture & Rural Development
Private Mail Bag 040
Port Vila
Vanuatu
Tel: (678) 22525
Fax: (678) 25265
Email: flehi@hotmail.com
Other CROP agencies
Forum Secretariat
University of the South Pacific
SPREP
Pacific biodiversity
Biodiversity hotspots
Breadfruit Institute
Hawaiian native plants
Intellectual property rights
Nature Conservancy
PBIF
PestNet
SIDS
WWF South Pacific Program
Other Pacific organizations
Foundation of the Peoples of the South Pacific
Micronesian Seminar
Te Puna web directory
Pacific news
Cafe Pacific
CocoNET Wireless
Island Directory
Pacific Islands News
Pacific Islands Report
Pacific Islands Travel
Pacific Time
South Pacific travel
Time Pacific
Interested in GIS?
DIVA-GIS
Tuesday, October 24, 2006
Seed plants of Fiji: an ecological analysis
Biological Journal of the Linnean Society
Volume 89 Issue 3 Page 407
by MICHAEL HEADS
An annotated list of indigenous Fijian seed plant genera is presented and comprises 484 genera and 1315 species in 137 families. The relative diversity of the largest families and genera in Fiji is indicated and compared with floras in New Caledonia and the Upper Watut Valley, Papua New Guinea. Differences and similarities appear to be due to biogeographical/phylogenetic factors rather than ecological differences or means of dispersal. Generic diversity for the seed plants as a whole is greatest between 0–100 m and decreases monotonically with altitude. However, in the largest family, Orchidaceae, maximum diversity occurs between 200–400 m. Fifty percent of the families are recorded from shore habitat. Twenty-seven percent of the families and 80 species occur in or around mangrove, where the most diverse families are Orchidaceae, Rubiaceae, and the legumes. Some of the mangrove-associate species are pantropical or Indo-Pacific but most are locally or regionally endemic. Fifty-six percent of the Fijian families are recorded on limestone. Twenty-nine species are restricted to limestone and 12 species usually occur on limestone. The importance of calcium in reducing the effects of salinity is emphasized and 39 species are recorded from both mangrove and limestone. A plagiotropic habit occurs in 38 species which occur on limestone or around beaches, and 20 of these are Pacific endemics. Genera restricted to higher altitudes include many present elsewhere in Melanesia but absent from Australia despite suitable habitat there, again indicating the importance of biogeographical and historical factors. Altitudinal anomalies in Fiji taxa are cited and include 7 anomalously high records from northern Viti Levu, a site of major uplift, and 22 anomalously low altitudinal records in the Lau Group, a site of subsidence. It is suggested that the Fijian flora has not been derived from immigrants from Asia, but has evolved more or less in situ. Taxa would have survived as metapopulations on the individually ephemeral volcanic islands always found at oceanic subduction zones and hot spots, and the atolls which characterize areas of subsidence. The complex geology of Fiji is determined by its position between two subduction zones of opposite polarity, the Vanuatu and Tonga Trenches, in what is currently a region of transform faulting. The large islands comprise fragments of island arcs that have amalgamated and welded together. There has been considerable uplift as well as subsidence in the islands and it is suggested that both these processes have had drastic effects on the altitudinal range of the taxa. Limestone and mangrove floras could have provided a widespread, diverse ancestral species pool from which freshwater swamp forest, lowland rainforest, dry forest, secondary forest, thickets, and montane forest have been derived during phases of uplift.
© 2006 The Linnean Society of London, Biological Journal of the Linnean Society, 2006, 89, 407–431.
Archive
RSS Feed
Alternative feed
Contact Tevita
Something new:
Agrobiodiversity Weblog: For discussions of conservation and sustainable use of the genetic resources of crops, livestock and their wild relatives.
